


How the Turn of the Century Turned Evolution Upside-Down
It was once common to hear evolution described as a unidirectional march of progress toward ever-increasing complexity, with humans, a recent species, being one of the most complex creatures that have ever lived (as remarked in dialogue in the 2000 film Hollow Man), if not the inevitable, perhaps even pre-determined, outcome of “the evolutionary process” (as depicted in the 2000 film Mission to Mars and the 1993 Star Trek TNG episode, “The Chase”). While such a description of evolution would cause migraines for virtually any biologist alive today, long, long ago, even a great many evolutionary scientists, including the likes of Darwin and Huxley, seemed to regard evolution as roughly synonymous with increasing morphological complexity over time, either resulting from some sort of internal drive toward complexity, as in orthogenetic theory, or because more complicated organisms were somehow just inherently more fit (the position that Darwin himself ended up taking, though not without reservation). Creatures who appeared to have obviously lost or reduced traits over time, such as parasites, were once heralded as examples of “anti-evolution” or “regressive evolution”. That evolution was inherently progressive and progress meant becoming more complex was simply taken for granted. So, what changed?
If you were a member of the scientifically literate public in the 1990s, had a subscription to National Geographic or Scientific American, frequented the Talk.Origins usenet group and/or spent hours browsing their online archives or any of the host of other online educational resources springing up around that time (both before and in the infancy of Wikipedia), it likely seemed this version of evolutionary theory was being violently overturned. The classic “five-kingdoms” system, in which the highest-level categories for life on Earth were the animals, plants, fungi, protists, and bacteria, was being supplanted by “tripartite tree of life” (Woese & Fox, 1997; Woese, 1990), which merged the animal, plant, fungus, and protist kingdoms into the domain eucarya, while splitting the bacterial kingdom monera into two domains: the bacteria and the archaea (Woese, 1990). Suggestions were being made that prokaryotes (the aforementioned bacteria and archaea) might be reductionist lifeforms who’d evolved to become simpler and more streamlined over time (Doolyttle & Brown, 1994; Lynch, 2006), and that an even more extreme version of this reductive evolution was responsible for the existence of at least some viruses (Topley & Wilson, &al, 1998; Nasir &al., 2012; Koonin & Wolf, 2012). The Rare Earth hypothesis, highly publicized around this time, hinged on the idea that only under unusually ideal conditions would complex, obligately multicellular life evolve. Taken together, these ideas seemed to suggest that such morphologically complex organisms were something of an evolutionary side-effect of microbial evolution (Carroll, 2001), and that nothing like the lifeforms most familiar to us would’ve had the chance to evolve if selective pressure had been much greater than it was. By the dawning of the 21st century, it seemed this was even beginning to trickle down into science fiction, as depicted in the climax of the 2001 film Evolution, in which we learn, as one character remarks, “the ‘simplest’ organisms are often the most fit,” as a gargantuan amoeba rampages across the landscape.
Of course, much of this was merely a sort of “delayed reaction”, culminating from changes that had already taken place in the scientific community finally filtering down into the science-consumer’s consciousness as years of information from articles and books became available on the newly-formed public forum that was the worldwide web. The progressive model of evolution was always contentious in scientific circles, even when it was the majority view (McShea, 1991), and had already received its first major blow from Ernst Mayr proclaiming that orthogenesis implied "some supernatural force" in a 1948 issue of Nature (Ruse, 1996), though progressivist assumptions would remain firmly entrenched until at least 1991 (McShea). However, that’s not to say this didn’t reflect events taking place in the scientific community at the time: Daniel McShea famously launched his inquiry into the subject of biological complexity at this time (1991, 1994, & 1996).
The biggest change during this period was the Woesean Revolution, named for the research of the eponymous Carl Woese, came in 1977, demonstrating that many of the creatures previously regarded as types of bacteria were actually far more closely related to eukaryotes (Woese & Fox, 1977; Woese, 1990), resulting in the aforesaid tripartite tree of life that has since become the standard in textbooks.
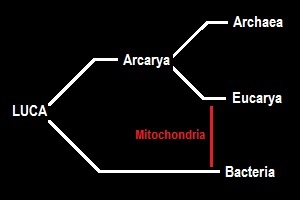
After Woese introduced the 3-domain system, this was quickly challenged by the eocyte hypothesis (Lake &al., 1988), which posited that the eucarya should nest within the archaea. This was followed by refinements/modifications like the TACK hypothesis (Guy & Ettema, 2011) and more recently the Asgardian hypothesis (Eme &al., 2017), each nesting the eucarya more deeply within the archaea. That we eukaryotes might actually be little more than an insignificant twig on the family tree of the archaea, who were once wrongly regarded as a type of bacteria, has quite literally flipped our image of life on this planet completely upside-down.
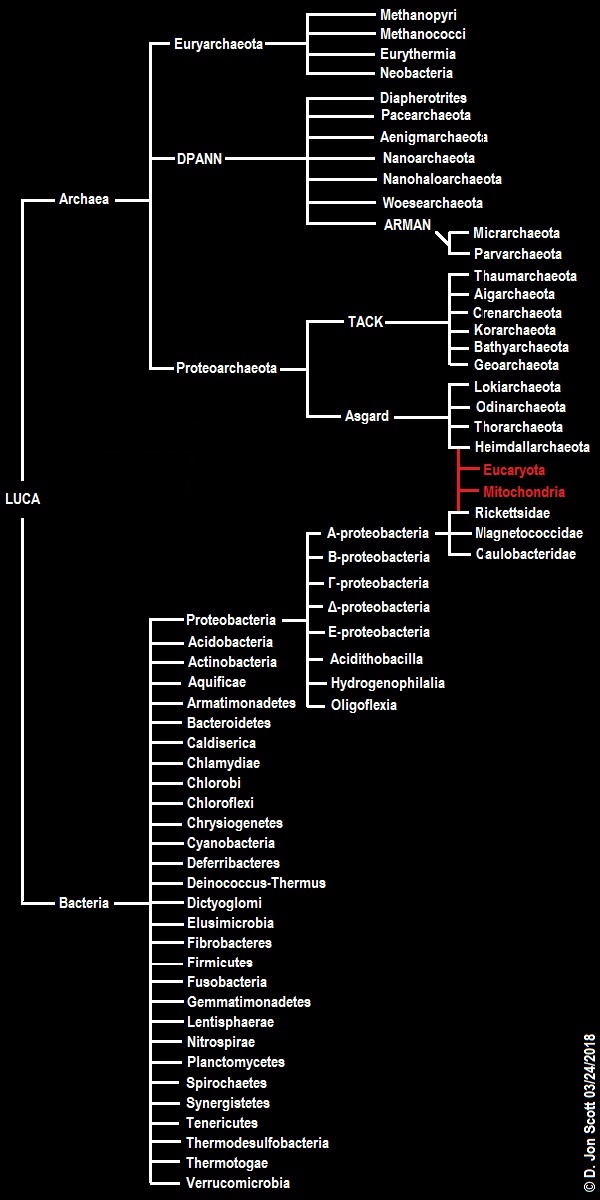
Today, “Eukaryotes-first” or “Eukaryotes-early” hypotheses, such as the nuclear compartment commonality or “NuCom” hypothesis (Staley, 2013), suggest that our “complex” form of life is actually the more archaic. This would consequently mean that most of the evolution that has taken place since our common ancestor with the bacteria has served to refine and simplify, resulting in what we now know as the bacteria and the non-eukaryotic archaea. The earlier karyogenic proto-coenocyte hypothesis (Egel, 2011) took this even further, positing that the primordial lifeforms from which the bacteria, archaea and eucarya descended may have been aggregate lifeforms comprising multinucleate, syncytial “super cells” far larger and more complex than anything alive today. Both hypotheses involve primordial life being essentially communal in nature, eerily reminiscent of the “pre-Darwinian” period of evolution endorsed by Carl Woese (Woese, 1990; Koonin, 2014).
While it is likely that some or all of these more recent hypotheses will fail to stand the test of time, it’s clear that the changes in evolutionary thinking that took place in the late 20th century have opened up a universe of new avenues of scientific inquiry. With even the long-cherished belief that primordial life must’ve been simple compared with modern lifeforms now being openly questioned as possibly nothing more than an anthropocentrism (McShea, 1991), it now seems more difficult than ever to envision what new discoveries or perspectives tomorrow might bring.
6
References
Carroll, S. B. (2001). Chance and necessity: the evolution of morphological complexity and diversity. Nature 409, 1102-1109 (22 February 2001). doi:10.1038/35059227
Doolittle, W. F.; Brown, J. R. (1994). Tempo, mode, the progenote, and the universal root. PNAS July 19, 1994 91 (15) 6721-6728
Egel, R. (2012). Primal Eukaryogenesis: On the Communal Nature of Precellular States, Ancestral to Modern Life. Life 2012, 2(1), 170-212; Received: 8 November 2011; in revised form: 29 December 2011 / Accepted: 11 January 2012 / Published: 23 January 2012 doi:10.3390/life2010170
Eme, L.; Anja Spang, Jonathan Lombard, Courtney W. Stairs & Thijs J. G. Ettema. Archaea and the origin of eukaryotes. Nature Reviews Microbiology volume 15, pages 711-723. (2017) doi:10.1038/nrmicro.2017.133 Published: 10 November 2017, Corrigendum: 27 November 2017, Corrected online 27 November 2017
Guy L1, Ettema TJ. The archaeal 'TACK' superphylum and the origin of eukaryotes. Trends Microbiol. 2011 Dec;19(12):580-7 Epub 2011 Oct 20. doi: 10.1016/j.tim.2011.09.002
Lynch, M (2006). Streamlining and Simplification of Microbial Genome Architecture. Annual Review of Microbiology Vol. 60:327-349 (Volume publication date 13 October 2006)
Koonin, E (2014). Carl Woese's vision of cellular evolution and the domains of life. RNA Biology. v.11(3); 2014 Mar 1 PMC 4008548, 2014 Mar 1; 11(3): 197-204. Published online 2014 Jan 16. doi: 10.4161/rna.27673
Koonin, E.; Wolf, Y. (2012). Evolution of microbes and viruses: a paradigm shift in evolutionary biology? Front Cell Infect Microbiol. 2012; 2: 119. Published online 2012 Sep 13. doi: 10.3389/fcimb.2012.00119
McShea, D.W. Complexity and evolution: What everybody knows. Biology & Philosophy. Volume 6, Issue 3, pp 303-324 (1991). https://doi.org/10.1007/BF00132234
McShea, D. W. (1994) Mechanisms of Large-Scale Evolutionary Trends. International Journal of Organic Evolution, The Society for the Study of Evolution. Vol. 48 December,1994 No.6 Evolution, 48(6), 1994, pp. 477-492
McShea, D. W. (1996) Perspective - Metazoan Complexity And Evolution: Is There A Trend? International Journal of Organic Evolution, The Society for the Study of Evolution. Vol. 50 April,1996 No.2 Evolution, 50(2), 1996, pp. 477-492
Staley, T. (2013). The Nuclear Compartment Commonality Hypothesis, Enucleation and the Evolution of the Bacteria and Eukarya. Journal of Astrobiology & Outreach, 1:3 2013. DOI: 10.4172/2332-2519.1000105
Topley, W.W.C.; Wilson, Graham K.; Collier, L.H.; Balows, Albert (1998). Topley & Wilson's microbiology and microbial infections.
Woese, C.; Kandler, O.; Wheelis, Mark (1990) Proc. Nati. Acad. Sci. USA Vol. 87, pp. 4576-4579, June 1990 Evolution Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eukarya. Contributed by Carl R. Woese, March 26, 1990
C R Woese and G E Fox, (1977). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Nati. Acad. Sci. USA